
The most recent Nobel Prize in Physiology or Medicine was shared among John O’Keefe, May-Britt Moser, and Edvard Moser for their work on how animals know where in the world they are, and what cells in the brain contribute to this inner map. Interestingly, the story has just begun - their longstanding views of how these neurons work are not the whole story, and there are many exciting and unresolved puzzles.
I had the pleasure of conversing with Dr. Mayank Mehta, a professor at UCLA, about his discoveries in the field of neurophysics at the interface of physics and neuroscience. His research shows that how we determine our place in the world is actually more complex than previously thought, and has solved several paradoxes exposed by the conventional theories. His excitement was contagious – especially when talking about his cool technology: virtual reality for rats!
Defining some terminology, and what the Nobel Prize was all about
D: I was reading your paper, and one of the things that first struck me was the amount of jargon in neuroscience that I’m not familiar with. So maybe we could start just by defining some of these things, like spatial selectivity.
M: Sure! So what was known, and what was part of the Nobel Prize that was given out, was the discovery of these so-called “place cells.” All animals move in space. And they must have a good idea of where they are; otherwise, they’re wasting energy. So how do they generate this perception of space? Long ago there was this idea called the cognitive map, which is a mental map of the world. People had done experiments to show that rats can have a map of the world, so they looked into where the map is in the brain. John O’Keefe and his colleagues way back in 1976 found that if you listen to one single neuron out of millions, one at a time, in the brain region called the hippocampus while the rat is just wandering around, that neuron will fire based on where the rat is in space.
D: I see. So in a different location, a different neuron will fire?
M: That’s exactly right. At one location one neuron will fire, at a different location a different neuron. And overall across all neurons you’ll have pretty much different neurons at all different places. That is the mental map or cognitive map. And these neurons have “spatial selectivity.” Now the question becomes: what is the nature of this map?
Map based on visual cues?
One kind of map is called an allocentric map which means a neuron fires at some place in the world regardless of what the subject is doing but dependent on the subject’s coordinates in the world. For example, the neuron will fire if the rat is two meters from the door and four meters from the kitchen. The things relative to which the rat’s position is defined are called the distal visual cues: the door is a distal visual cue, the kitchen is, and so on. Such a cognitive map is called an allocentric map because it is with respect to things outside the body. So the idea is that the way these neurons in the hippocampus localize and fire in a given place is determined by the distance from various distal visual cues or landmarks. And to some extent there is evidence for that. If you put the rat in a box and you put different marks on the walls – one wall is plus sign, others are circle, triangle, or wave pattern – and a neuron fires at the 12 o’clock position two meters from the corner, if you rotate the box the place cells rotate with it.
D: I see. So it’s not the absolute coordinates at that point, it’s relative to the cues.
M: Exactly.

Source: simplecodeworks.com/hypermaze/index.html
Map based on multi-sensory cues?
But then – the puzzle is that it’s also known that if you turn off the light, there are no visual cues but the neuron will more or less fire at the same place it was firing before with respect to the cues! So what the hell happened? One possibility was that if the place cells continue to fire when it is in darkness, maybe it’s driven by other cues like smell, sound, or texture. And to some extent that was supported experimentally when they took blind rats, and it turns out they have as good place cells as sighted rats! No difference at all. So place cells are more complicated, and are not determined by just visual cues as people thought. They are made up of multiple sensory cues. So that’s the multi-sensory hypothesis we propose.
Map based on self-sensing cues?
The story gets more interesting: where the place cell fires might not just depend on the stuff outside in the world, like smell and sound but also on what he himself is doing. Like maybe the rat’s brain is keeping track of how fast he is going in the forward direction, how many left turns he makes, and so on. If he was doing that he could still localize and tell where he is. So with that signal – the number of right and left turns – he’s making a path in the vector sense. And in order to do that you need a cue called the vestibular cue. There’s an organ in our inner ear that’s used for the sense of balance and head rotation. There’s a canal that’s filled with fluid called the vestibular apparatus. And this is called the vestibular signal, which tells you how much you have turned or accelerated.
So now we’ve got all the players in mind. So before, one idea was that place cells are governed by distal visual landmarks. Instead, that doesn’t seem to be just happening. It does partially happen – when you rotate the landmarks the place cells rotate – but five or six other things can contribute. Sound, smell, texture, locomotion, memory, reward – everything is contributing. So now the problem gets really complicated. What is a place? All these experiments are really hard to do in the real world, because when you do an experiment, everything is changing and you have no control over it to manipulate it. So that’s where we said: why don’t we develop a virtual reality?
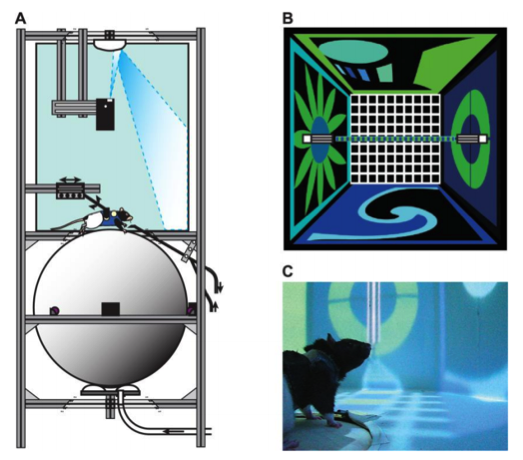
Source: Ravassard P, Kees A, Willers B, et al. Multisensory control of hippocampal spatiotemporal selectivity. Science. 2013;340(6138):1342-6.
Making a virtual reality: can visual + vestibular cues alone create a map?
In this virtual reality, a little tuxedo is put around the rat’s chest, and at the back of the tuxedo or harness there is a little Velcro, and the Velcro holds the rat in one place. Now he happily sits there wearing a tuxedo and he’s on a big floating ball and as soon as he starts to walk on the ball, the Velcro holds him in one place but the ball is calibrated to have the same weight as the rat and starts to rotate. The rotation of the ball is picked up by a bunch of computer mice. The mice pick up the signal of movement, and the computer creates a virtual world with all sorts of visual and auditory cues that we as the experimenters have decided, and the computer updates an image to a projector that is above the rat’s head. Through some more gimmickry, it projects an image not just in front of the rat but all around it: behind him, above him, all the way up to his feet so he can see his shadow change.
D: Wow, that’s very detailed.
M: Yeah, exactly, because we want to be very careful. If we just put a screen in front of him – rats are not like us, they don’t know what a screen is. He’d think this is bogus! Even IMAX is not [convincing], because maybe they’re checking out the floor all the time. We made sure that it goes all the way to the tip of his nose. Only half an inch area around him is not covered, and that much we can’t do, but that he probably can’t see anyway because his eyes are on the sides.
D: That’s incredible technology!
M: Yes, the credit goes to the brilliant and hard work done by the lab members. We put the whole virtual reality rig inside a dark room so no other cues can come in and no humans can come disturb it. The room is sealed up and there is a 60-decibel sound barrier so the smells and sounds and light – nothing from the external world can go in. And then we ask him to walk in this world and see if he buys into it. We did a lot of experiments to make sure he does!
So we wanted to see how the rats form maps. In the virtual world the rat is put on a table. We ‘release’ him from one of the corners and we have a tiny unmarked reward zone. If he wanders around and reaches the zone, he gets sugar water from the reward tube in front of him. Then, while he’s drinking the reward we ‘teleport’ him to a different part of the table and again he has to find the unmarked reward zone. So now he’s not just going based on a scent trail or something like he could do in the real world – in the virtual world he can’t do that. So this would 100% for the first time show that if the rat goes somewhere in the virtual world, he is going there only because of adding up vectors with respect to landmarks. And he can do it!
Challenging the status quo: a new model for “place cells”
All right, now we know that he can form a map – but what do the neurons do? So we said, let’s simplify it and make it a one-dimensional world. He just runs back and forth on a simple two-meter-long track. He reaches the other end, gets a little reward, turns around, and runs back. Just to be careful, we made a real room next door in the lab just like the virtual room with a real track. We made the same rat run in the real world and virtual world. We measured the same neurons in both worlds while he was just running back and forth. And this gets to a very deep question – what is reality?
Let’s compare the neurons as he runs in identical real and virtual worlds. Until that point, people had the theory that there should be no neuronal difference between the real and virtual world because the landmarks are the same and the rat's behavior is quite similar. But that’s where we had our first major surprise, which no one expected. What we found was that the very fact that the rat was running in the virtual world resulted in 60% of the space-mapping brain shutting down. 60%! That’s a huge thing. We and everyone else expected there’d be no difference! The rat himself was behaving perfectly well in the virtual world; he was running from the beginning, he accelerated, and even though he had no need to decelerate when he reached the end of the virtual world (unlike in the real world where if he doesn't decelerate he’ll hit his nose at the end of the table) he knew that he was reaching the end and slowed down. The rat knew exactly where he was and what he was doing. And still 60% of neurons simply shut down.
Then we looked at neurons that are actually active, the other 40%. Turns out, of these 40%, some neurons are active when the rat is running [head-first] in both forward and backward directions. Other neurons only fire in one direction – that much we also know to be true in the real world. Half of the neurons are unidirectional and half are bidirectional. Now what has been known in the real world is that if a neuron is active in both directions, that neuron will fire at the same place in both directions. If he was going forward on a 200cm-long track and in the forward direction the neuron fired after 20cm from the starting point, in the reverse direction the neuron will now fire after 180cm from the other starting point.
D: So it’s the same absolute spot.
M: Exactly right, it’s the same absolute spot. And this we verified. When the same rat ran in the real world we indeed found that half the neurons fire in one direction and the other half fire in both directions and invariably fire at the same spot. Then we looked at the virtual world. Just like the real world, half the neurons fire in only one direction. And the other half fire in both directions which was pretty much exactly the same. But then when we looked at these neurons firing in both directions, there was a second major surprise. These bidirectional neurons actually never fire at the same absolute position. Never.
D: Wow, that’s crazy.
M: Yeah, it’s totally crazy! But it was not random either. These neurons were firing actually at the same distance in both movement directions. Which means that if the rat started to run and the neuron fired at 20cm going out, then when he starts to run back, the neuron again fires within 20cm on his backward journey.
D: I see. So by definition, it could never be at the same spot.
M: Exactly. Because they are keeping track of the same distance not the same place, and the distance will be diametrically opposite. So now what we find is really funny. The only two things that were telling the rat where he is, the distal landmarks and the locomotion cues … instead of giving beautiful place cells, these cells are actually counting distances [in virtual reality]. So that was second major discovery as far as I think, because the classic idea is being challenged.
D: Right. What I find so incredible is that whether they’re in the real world or virtual world they’re still behaving the same, it’s just the neurons that are doing something different.
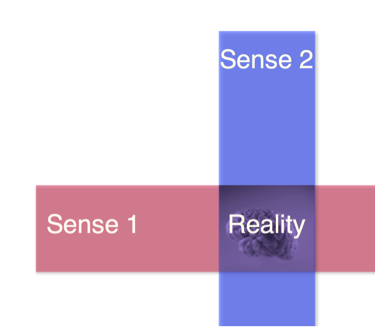
M: Exactly, that’s right! The rat’s behavior is [the same] but the neurons are doing diametrically opposite things. And 60% of total neuron function is down. The standard cognitive map [model] cannot explain this. We have a new theory, which we call the multisensory pairing hypothesis. The idea is that where a hippocampal neuron will fire is determined by a combination of more than one sensory modality, which are consistent with each other. And that in our definition is reality. When we say something is real, it is real when all the sensory modalities are consistent with each other all the time.
D: Interesting. So could you give an example of that? What would be real and not real?
M: When the rat is running in the real world there may be a little scratch on the table. As result what happens is that whenever the rat is running in the maze, because the little scratch on the ground is fixed, the neuron responds to a combination of the scratch and the distal visual cue and the neuron will fire in the same place. Because two different things are coming together and are consistent. In the virtual world, the ball he is running on has some scratches but the same scratch doesn’t appear in the same virtual place all the time... So that’s what we think is going on and this is something nobody expected. We feel this combination of multiple modalities is what the hippocampus [centralizes]. And that starts to actually make sense if you look at biology. If you look at other parts of the brain in the neocortex – you have the visual cortex where neurons only care [about] visual cues, auditory cortex for sounds, and olfactory cortex for smells. The question becomes, who the hell will put all this together? You don’t just smell things or see things, you have it all together and we believe that’s the job of the hippocampus. If you look at brain anatomy you see all these different regions sending their input to the hippocampus, so we think that’s what it’s doing. That’s what space is – putting it all together. In the virtual world, the one-dimensional locomotion cues and the visual cues are in register, and that consistency gives a [map of, or sensation of] distance. (Because when the rat starts to run in the forward direction he is accelerating and when he reaches the end he is decelerating.) But when he comes back, in the same place now where he was accelerating on the track, now he is decelerating. Now his internal movement is not consistent and that is, according to our theory, the reason why the neuron cannot fire in the same place. But on the other hand when he is leaving the beginning of the track the kind of optics that are generated will be exactly the same in both directions. And as a result the neuron will now fire at the same distance, with the world rushing by at the same rate.
Improving the multisensory pairing model
We wanted to check whether the neurons in the rat can form a map of the world using only distal landmarks. In this one-dimensional path, not only the landmarks but [also] locomotion cues are telling him where he is. So we need to get rid of that by designing a task where the only thing that tells him where he is are the visual landmarks. Nothing else. In order to do that we have to put the rat in a two-dimensional maze. We have to make him walk around randomly so whenever he’s in the same place he never visits it with the same acceleration. So we designed a virtual maze and real maze where we threw rewards randomly. As a result, he is accelerating and decelerating randomly at any place. We put him in the virtual world, and the neuron fired totally randomly. All over the place! That was consistent with our idea, since the multisensory idea says that in order to form a map of the world, multiple modalities have to be consistent. In one dimension, the acceleration was consistent with visual cues, but in two dimensions the acceleration is not consistent, so there is no selectivity. That challenges the [Nobel-winning] idea that with only distal visual cues you can add up those distances and generate a sense of place.
D: So in the multisensory model, do all of the cues have to line up, or just more than one?
M: In our current thinking – who knows, we are probably wrong too--but we think you need at least 2 different modalities. But they have to come together repeatedly in the same configuration or the neuron won’t fire very well.
D: Just to clarify, the place cells which were previously thought to explained by distances from landmarks, you’ve now replaced with the multisensory pairing model.
M: Yeah, we think the multisensory pairing model might do a better job in explaining many results. Instead of the hippocampus computing a map of the world, we suggested that that it will actually do multisensory paring and that place cells are just one manifestation of it. The [multisensory] pairing explains some paradoxes that people see in the real world, like how place cells persist in darkness, and blind rats have place cells, and place cells become distance cells, and so on.
The future of mapping reality
D: So obviously you’ve been working on this research for a long time and you wrote multiple papers on it. Have you found that in this field people naturally gravitate into two camps of thought – multisensory modality vs place cells, or is it sort of large scale discussion?
M: In a way, I would not say that lots of people buy into the multisensory pairing hypothesis. It’s new. It will still take some time to be fully tested and change the worldview. I think it will happen at some stage; so far people are thinking about it and checking this and that, so we will see.
D: Well you’ve convinced me!
M: {Laughs} Well, we think the results are compelling. But that’s how science works. There are some ideas that are established for a long time and something comes and challenges it and even If it does a better job, people will be reluctant for a while.
D: That’s definitely true; I think we see that in pretty much every field. In your opinion, what do you think is the best way in general for science to handle new viewpoints that challenge the status quo?
M: I think in the end it’s good for science to be careful and not change the worldview every single day…it’s good to be cautious. But at the same time, in the end, we have to remember that science is also done by scientists, who are human beings. They have memories, some formed ideas that they like, perhaps which they came up with or believed in for many years, so even if the bare raw facts can be pointing the other way we are all human beings and are all a little bit biased.
D: {Laughs}
M: {Laughs} I certainly am, I don’t know about the others. So I think I understand that and I don’t know how to change that – it’s just the way the world is. I can hardly change myself, let alone the rest of the world. The only place I think it starts to make a difference is when some surprising thing has been demonstrated overwhelmingly and people still don’t take it into account or ignore it or dismiss it, or say it’s either obvious or wrong, then science is not going in the right direction. But right now that’s not happening with this; it’s still pretty new. In time we’ll see where this goes. And in the meantime we’re busy writing up other things. This is only part of the surprises!
D: {Laughs} Fair enough, I’ll ask you again sometime. Thank you so much! This has been so interesting - I didn’t know about any of this and it’s really exciting to hear about.
M: Yeah, it’s a fascinating subject, isn’t it?
D: Absolutely!
Appendix: additional mysteries in the brain
M: We started looking more carefully at the kind of activity in the brain when the rat was running randomly. Was there any pattern? And there we found another surprise: when the rat was running randomly, even though the overall activity of neurons were random and cells were firing everywhere, whenever the cell was active it was not just active in a punctate fashion (like 1 or 2 spikes and then over). It didn’t look like that. Whenever the neuron was active it would be active for a long period of time, like 2 seconds long. Sometimes 4 seconds and sometimes half a second. Different neurons are active for different amounts of time. If you look at the same neuron in the real world and virtual world, the neuron will be active for same duration in the real and virtual worlds. Within the 4 second long burst there is a very exquisite precise structure which is a function of oscillations generated by the brain and even that was unchanged. So based on this we have now the next level of theory: we think that in addition to this multisensory pairing that generates place, our hippocampus is hardwired so that whenever the hippocampus is active it will be active for more or less a couple seconds. Different neurons have “memories” – from half a sec long to 5 or 100 seconds long. The reason we think the brain is doing this is something we take for granted in physics all the time. Whenever anyone starts doing anything in physics the first thing we do is talk about continuity equations. We start with the assumption that space is continuous. That’s a big deal, and it’s continuous in the virtual world as well. Imagine you’re in one place and simply move your head by 10cm. It’s a tiny amount. The world really doesn’t change that much. If you now try to recalculate where you are, it will be a tremendous waste of energy to recalculate every millisecond let alone 100 milliseconds. So in order to save energy, in evolution the brain has figured out a way out by saying that if you’re somewhere and you’re going around with some average speed, if there’s some neuron that’s active then for the next couple seconds it’s more or less the same. That's we think is going on. That’s what we call a motif.
Here’s another story: It’s well known that when we start moving in space, in addition to the place cells that appear in the hippocampus actually one more funny thing starts to happen. If you look at the entire hippocampus, the activity becomes rhythmic in time. And the rhythm is about 8Hz, 8 times a second. That rhythm appears whenever a rat is running, and when he stops the rhythm it goes away. It reappears when he’s dreaming in the REM sleep. The 8hz rhythm appears in the real world and virtual world. But there is something even more funny. When he runs in the real world, as he runs faster the rhythm starts to get faster. So if he’s running 10cm/sec the rhythm is 8 times a second, if he’s running at 100cm/second the rhythm becomes say 9 times a second. So how come the rhythm has something to do with running speed? It’s a long story but we believe it has something to do with space, time, and memory and have a couple papers on it. But the most surprising thing that I never ever expected is that when he starts to run in the virtual world, the 8hz slows down to 7hz, and as he runs faster and faster the rhythm doesn’t change at all. It’s fixed at 7hz. So when the rhythm slows down its frequency is decoupled from running speed in the virtual world – a little mystery.
D: So in that case – we were talking earlier about the duration of the neuron firing. When you’re running faster and the 8z is changing, does the duration change also? Because now in a shorter amount of time your world is changing more than it was before.
M: That’s a great question, and in order to give you a full answer it will take a long time. I’m simply going to say that as far as single neurons are concerned, the duration for which a neuron is active doesn’t change. But that happens in a very funny way which is complicated. It’s a little long so I’ll pretend you didn’t ask that. {Laughs}
Aghajan et al. (2015) Impaired spatial selectivity and intact phase precession in two-dimensional virtual reality. Nature Neuroscience 18, 121–128.
O'Keefe, J., and Dostrovsky, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely‐moving rat. Brain Research 34, 171-175.
Ravassard et al. (2013). Multisensory Control of Hippocampal Spatiotemporal Selectivity. Science 340, 1342-1346



